
研究内容
オタマボヤは「最も単純な脊索動物」
私達は、ワカレオタマボヤ (Oikopleura dioica)という脊索動物を材料にして、発生や体づくりの研究を進めています。
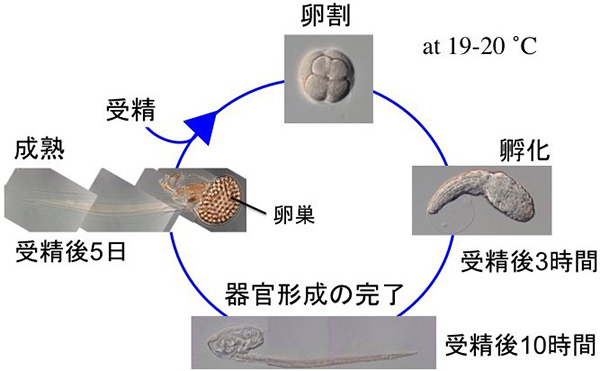
図はその生活史です。オタマボヤは脊索動物門(一生のどこかでオタマジャクシ型の体になる動物群。私たち脊椎動物も含む)に属しますが、体の細胞数が4500個あまりと少ないです。受精後3時間でオタマジャクシ幼生が孵化すること、10時間には一生分の細胞分裂が終わり大人と同じ体になること、ゲノムサイズが56-70 Mbあまりと小さいこと (寄生性の動物以外では最小)、そしてわずか5日間で次世代になるなどの特徴があります(20°で飼育した場合)。
このようにオタマボヤは、「線虫のように単純な体の実験動物の利点」と「脊椎動物と共通したオタマジャクシ型の発生」とを兼ね備えた生物といえます。
この実験動物としてのポテンシャルに着目して、私達はこれまでに
(1) 遺伝子の機能的スクリーニング (遺伝子の働きを調べる);
(2) 発生のライブイメージング(細胞の振る舞いを見る);
(3) ゲノム・トランスクリプトーム(遺伝子のリスト化);
(4) 組織学的手法や3D画像の取得方法(形をとらえる)
などの実験基盤を揃えてきました。これらをもとに、オタマボヤの生物学的な特質を活かした研究を進めています。以下、主な研究内容を2つ説明します。
(1) ハウスの建築
オタマボヤは体外に「ハウス」とよばれる、セルロースを含んだ袋を分泌して、その中に棲んでいます。
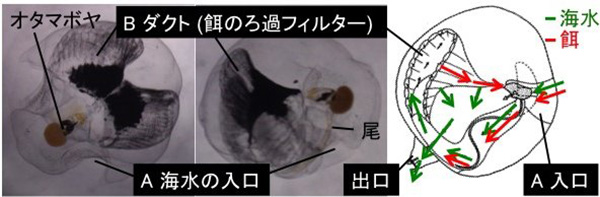
図に示すのは、ハウスの中に取り込まれた海水と餌の動きです。餌の代わりに墨汁を食べさせると、墨汁の粒子が取り込まれているのが分かります。
ハウスは単なる袋ではなく、海水中の餌をこし取り食べるための餌フィルターなどが3次元的に配置された、複雑な構造をもっています。
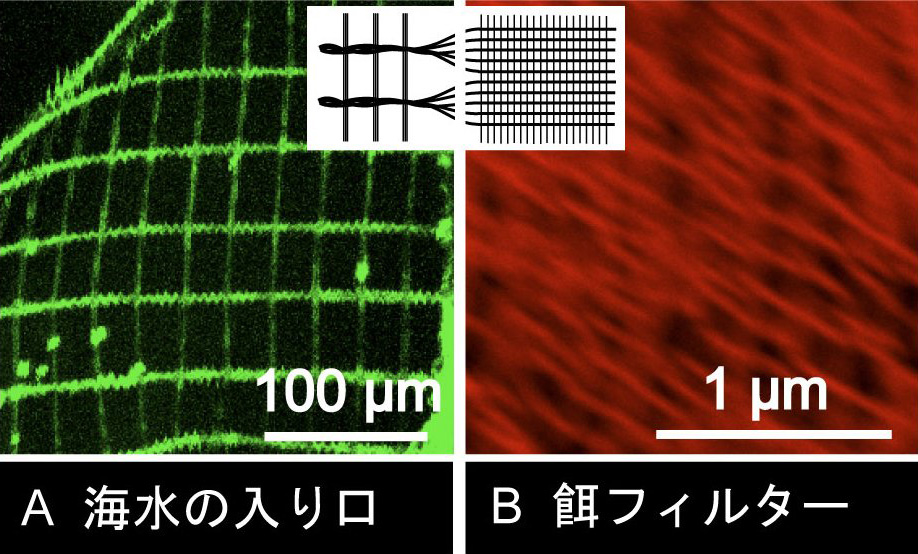
図はハウスの一部を高倍率でみた写真です。ハウスの各部位には、繊維でできた形の異なる網目があります。
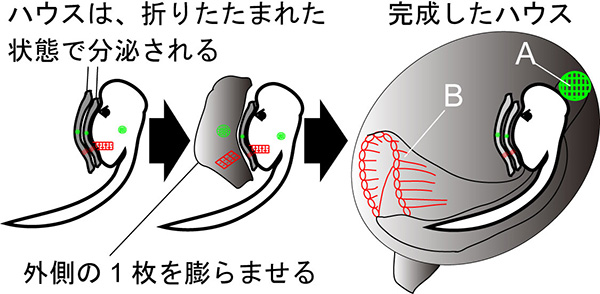
ハウスは「折りたたみ式」です。図に示すように、オタマボヤの体は、2-3枚のハウスを折りたたんだ状態(ハウス原基と呼ばれます)で体にまとっています。外側の一枚をふくらませると、数分以内にハウスが完成します。古くなったハウスは脱ぎ捨てられ、一日に5-8回、新しいハウスを作り直します。
ハウスを分泌する表皮細胞は特徴的な並び方をもっており、かつ、細胞数や位置に個体差がないことが分かっています (Kishi et al., 2017)。
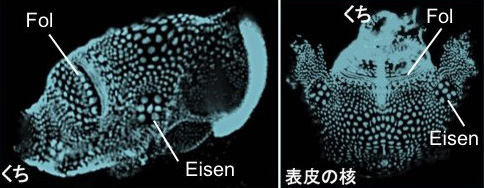
図の左は表皮細胞の核を側面からみたもの、右は「あじの開き」にしたものです。この表皮上で繊維を分泌して、ハウスの立体形状を設計すると考えられます。この「製造工程」について、以下3つのスケールで理解に取り組みます。
- 細胞によるセルロース繊維の合成(糸の合成、10-100 nm程度)
- セルロースの網目形成 (布の形成、1-10 µm)
- ハウスの形態(立体構造、数 mm)
ハウスの作り方は生物学の新しい研究領域といえます。「セルロース繊維の編み方により立体的な形状を決める」という点から、工学技術としての発展性も期待されます。生物学的な実験はもちろん、数学・物理・工学の専門家とも連携した数理モデルからもヒントを得て理解を試みています。
(2) 左右非対称形成
私たちの体は、直交する3つの軸をもちます(前後・背腹・左右)。左右ができるしくみは謎が多く、動物群ごとに異なるしくみが知られています。オタマボヤは一世紀も前に、初期胚に左右非対称性があることが知られていました。
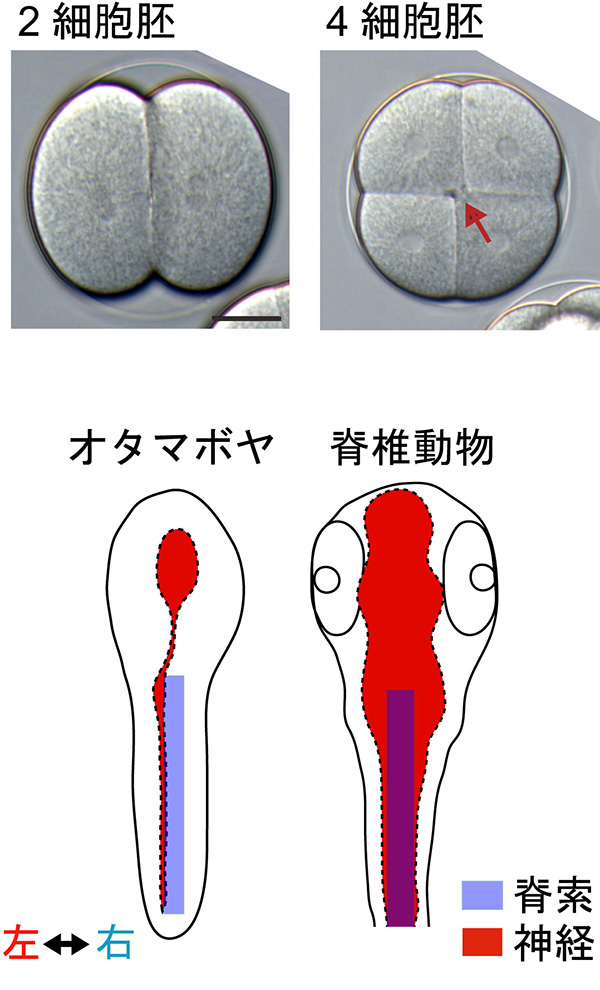
図の上は初期胚(植物極側)、下はオタマジャクシ幼生の模式図(背から見たもの)です。2-4細胞期にかけ、割球が個体差なく同じ向きにズレます。オタマジャクシの尻尾を見ると、神経の束(神経索)が約90°ずれ、(背側ではなく)左側を走っています。私達はこの左右形成のしくみが背腹をつくる経路を90°回転させたものである可能性を提唱しました (Onuma et al., 2020)。
この「最も単純な脊索動物」には、左右形成の新しい原理が眠っていました。今後は、「初期胚の細胞の位置がずれるしくみ」や「いつ、どのように右側と左側の遺伝子発現の差が生じるのか?」を調べていきます。これらを通じて、脊索動物の特徴であるオタマジャクシ型の発生を維持しながら、体づくりのしくみがどのように変わりうるのかという問題に迫りたいと考えています。
この2つに限らず、独自の発想やアプローチを大事にして、さまざまな研究テーマを開拓しています。試料や技術の提供を通じて、国内外の方々との共同研究を進めています。さらに、科学教育や啓蒙活動の素材としての普及を目指しています。ご質問等ありましたら、遠慮なくお知らせください。
これまで、卒業生が実施したテーマです
- オタマボヤの精巣特異的ヒストン遺伝子の配列決定
- オタマボヤ成体のSEM画像の取得 (Onuma et al., 2017)
- 可変色蛍光タンパク質Kaedeによる割球標識 (Kishi et al., 2017; Onuma et al., 2020; Morita et al., 2020)
- ゲノム・トランスクリプトーム情報の構築 (Wang et al., 2015; 2020)
- 動物初となる二本鎖DNAによる新規ノックダウン現象 (DNAi) の発見と、母性因子の大規模スクリーニング (Omotezako et al., 2013; 2015; 2017)
- 卵の減数分裂停止をになう母性因子PP2Aの発見 (Matsuo et al., 2020)
- オタマボヤの生殖細胞と生殖巣形成
- DNAiのしくみを、RNAiとの類似性から探る
- オタマボヤの体内を長距離移動する細胞の発見 (Kishi et al., 2014)
- 表皮のパターン形成の記録 (Kishi et al., 2017)
- レーザー照射による表皮細胞の破壊
- オタマボヤのY染色体 (Wang et al., 2020)
- 初期胚の細胞運命限定と胚性遺伝子発現 (in prep.)
- 初期胚に局在する母性mRNA (in prep.)
- 外胚葉と内胚葉の上皮間つなぎ変えによる口形成 (Morita et al., 2020)
- 孵化幼生の組織境界形成とカドヘリンの遺伝子発現
- 1-2細胞胚に形態レベルの左右非対称性は存在するか?
鹿児島大学は、錦江湾や奄美群島の水系にアクセスしやすく、ワカレオタマボヤ以外のオタマボヤの入手にも最適の立地にあります。学生のみなさんには、これらをフルに活用して、新しいチャレンジをして欲しいと思っています。
研究室の方針
各自、最低1つの研究テーマを担当します。教員をはじめ研究室メンバーと相談しながら、実験や解析を自ら行います。知的好奇心を形にするために必要なのは「正直さ(失敗を隠さない)」と「相談する(対話する)」です。これを通じて、実験の計画・準備・結果の解釈、学会発表、論文作成までを一通り経験します。大学院生も、筆頭著者で論文を書くことを目指します。
